
Vol. 35, n.º 4, 2002
|
REVISTA
ESPAÑOLA DE
Vol. 35, n.º 4, 2002 |
Fernando Reinoso Suárez
Catedrático Emérito de Anatomía y
Neurociencia. Departamento de Morfología. Facultad de Medicina. Universidad
Autónoma de Madrid.
Académico de número de la Real Academia Nacional de Medicina.
Las primeras noticias sobre D. Santiago Ramón y Cajal, las recibí de mi padre, médico rural en la Alpujarra granadina. Me figuro que fue con motivo de algún acontecimiento de relieve, posiblemente la muerte de D. Santiago en el año 1934. Me dijo que el hijo del médico de un pueblecito de Navarra había llegado a ser un sabio universal a base de trabajo ilusionado, bien hecho, bien acabado y dedicación intensa, lo recuerdo porque sobre estos motivos del éxito de Cajal me insistió muchas veces en años posteriores, seguramente con objetivos formativos. Mi padre había conocido a Cajal durante el período en el que hizo el doctorado en Madrid y le había impresionado su enorme personalidad. Creo que fue cuando me exigieron como texto de Histología en el primer año de Medicina la Histología de Cajal y Tello cuando mi padre me regaló, junto con este libro de texto, los libros de Cajal «Tónicos de la voluntad» y «Recuerdos de mi vida». Tengo que confesar que entonces leí sólo la primera parte de esta segunda obra «Mi infancia y juventud»; la segunda parte «Historia de mi labor científica» la leí años más tarde y llegué a conocerla en profundidad, lo que me permitió hacer con cierta facilidad la introducción a la edición que hizo de la misma Alianza Editorial en el año 1981 (1).
En el mes de julio pasado ha hecho cincuenta y dos años que comuniqué a mi maestro, el profesor Escolar, mi decisión de dedicarme a la investigación neurocientífica. Una vez que constató que mi decisión era firme me entregó los tres tomos de la «Histología del sistema nervioso del hombre y los vertebrados» de Cajal y me dijo «estudia esta obra en profundidad y habrás colocado un sólido fundamento a tu formación como científico». Aunque no fue sencillo, le hice caso y sigo estándole profundamente agradecido por el consejo. Pues, a mí me ha reportado considerables ventajas el conocer la obra de Cajal, y más concretamente el tomarla como punto de partida en mis investigaciones. Hay una anécdota por mí muy querida, por lo que la he repetido en numerosas ocasiones. El año 1959 fui invitado a dar una conferencia en el Instituto de Fisiología de Pisa que entonces dirigía el profesor Giuseppe Moruzzi. El tema era «El sistema reticular ascendente de activación», tema de moda de la época y que había sido definido en 1949 por el propio Moruzzi junto a Magoun. Era creencia común que el camino principal de este sistema hasta la corteza cerebral tenía como último escalón el núcleo reticular del tálamo. Yo afirmé en mi conferencia que de acuerdo con hallazgos anatómicos y funcionales de mi laboratorio esos impulsos ascendentes activadores parecían seguir esencialmente un camino subtalámico. Igualmente negué que el último escalón de ese camino fuese el núcleo reticular del tálamo, fundamentándolo también en hallazgos anatómicos y funcionales de mi laboratorio. En la discusión se me dijo que el papel del núcleo reticular del tálamo, como estación final de estos impulsos hacia la corteza cerebral había sido suficientemente demostrado tanto anatómica como fisiológicamente en una documentada monografía que firmaban un prestigioso anatómico y un prestigioso fisiólogo, y que había aparecido el año anterior a mi conferencia. Para defender mi punto de vista junto a los argumentos que figuraban en mis trabajos utilicé uno que resultó de mayor peso, y es que Cajal había demostrado que, imprevistamente, los axones de las células del núcleo reticular del tálamo, se dirigen no a la corteza cerebral, como lo hacen el resto de núcleos talámicos, sino hacia atrás y abajo (fig. 1). Cajal llega a definir este núcleo de estación centrífuga, puesto que sus células envían sus cilindroejes no a la corteza cerebral sino a la periferia, a núcleos por él desconocidos. Como siempre he hecho, había comprobado que nuestros resultados no contradecían hallazgos de Cajal, sino que por lo contrario tanto el camino subtalámico como la no participación del núcleo reticular en la proyección a corteza confirmaban descripciones de este. El matrimonio Scheibel confirmaba dos años más tarde la proyección a tálamo del núcleo reticular y hoy nadie piensa en el núcleo reticular del tálamo como la estación final en el camino a corteza de impulsos reticulares ascendentes. Igualmente, las nuevas técnicas han demostrado que los axones del núcleo reticular terminan en el tálamo y el mesencéfalo. Recientemente nuestro grupo ha observado que también lo hacen en el núcleo reticular oral del puente (2), que junto con tálamo y mesencéfalo, son las estructuras periféricas a que se refería Cajal.

Fig. 1.
Cuando verdaderamente tomé conciencia de la trascendencia de la obra científica de Cajal fue durante mi estancia posdoctoral en Alemania en los años 1951 y 1952. Estuve en el Neurophysiologische Abteilung del Max Planck Institute für Hirnforschung en Göttingen. Tanto su director, el profesor Kornmüller, como los componentes mayores del mismo eran grandes admiradores de Cajal. Para Kornmüller uno de los aspectos más importantes de la obra de Cajal, además de su extensión y fidelidad, era el sentido funcional que imprimía a todos sus hallazgos y la clara distinción que hacía siempre entre hechos demostrados y consideraciones sobre los mismos. Asistió con entusiasmo a la reunión científica que se celebró en Madrid el año 1952 con motivo del centenario del nacimiento de Cajal.
Durante esta estancia en Alemania conocí al profesor Hugo Spatz en cuyo Neuroanatomische Abteilun de Giessen tuve ocasión de pasar largas temporadas en el transcurso de los años cincuenta. La biblioteca de este Departamento estaba presidida por una enorme fotografía de Cajal. Para Spatz, Cajal era el fundador de la ciencia neurobiológica actual y afirmaba que mientras haya personas que quieran conocer el funcionamiento del cerebro, que será siempre, tendrán que apoyarse en Cajal.
También en el año 1952 escribió McCulloch refiriéndose a Cajal: «Su teoría neuronal ha sido tan fuertemente establecida como la base de nuestra ciencia, que nosotros ignoramos qué pasaba antes, y hemos olvidado que él fue el primero que la propuso». He traído esta cita por dos motivos: 1) para demostrar que no sólo en Alemania sino en todo ambiente neurocientífico Cajal era y es la primera figura de la Neurociencia, así como que 2) que si de la más trascendental aportación de Cajal a la ciencia, la teoría neuronal, nos hemos olvidado que fue Cajal su autor, lo mismo ha pasado con otros muchos hallazgos y teorías expuestas por Cajal a finales del siglo XIX y principio del XX. Es curioso que muchos de los que han utilizado y utilizan a Cajal como bandera política no conozcan qué hizo Cajal e incluso se olvidan de él cuando sería obligatorio citarlo.
Es una realidad la admiración de todos los neurocientíficos por Cajal. Yo lo he comprobado constantemente a lo largo de estos cincuenta últimos años. Podría poner como ejemplo a Nauta, el más prestigioso neuroanatómico de la segunda mitad del siglo XX, que apoyó en Cajal todas sus importantes contribuciones, o a Hubel, Premio Nóbel de Fisiología y Medicina, que escribía en 1978: «Su monumental Histología del Sistema Nervioso del Hombre y los Vertebrados es aún reconocida como el más importante trabajo publicado en todos los tiempos en Neurobiología». Sin embargo deseo hoy referirme a uno de los hombres que ha hecho más por la Neurociencia en los últimos treinta años y que desgraciadamente ha fallecido el pasado mes de junio a los 70 años de edad en su casa de Rockville, Maryland. Me refiero a William Maxwell Cowan al que quiero con este motivo rendirle un homenaje de cariño y respeto. El profesor Cowan dirigió importantes departamentos dedicados a la investigación Neurocientífica en los que formó numerosos discípulos que hoy ocupan destacados puestos de responsabilidad en la Neurociencia mundial. Fue editor de la revista Journal of Comparative Neurology y fue el fundador y primer editor de la revista Journal of Neuroscience. Presidente de la Society for Neuroscience y miembro de la Nacional Academy of Science, en los últimos años ha contribuido a promocionar la investigación científica como Director Científico del prestigioso Instituto Médico Howard Hughes. Max Cowan era un entusiasta admirador de Cajal, participó eficazmente en un intento de traducir la obra de Cajal «Histología del Sistema Nervioso del Hombre y los Vertebrados» al inglés, que tendría que presentarse como homenaje al sabio español en el centenario de su «año cumbre»: en 1988. En 1888 formuló Cajal la teoría neuronal. Algún día debería contarse la historia del fracaso de esta traducción debido a la incapacidad de unos y al egoísmo y visión corta de otros. Max Cowan visitó el Instituto Cajal en varias ocasiones y escribió mucho y bueno sobre Cajal. Yo me voy a limitar a reproducir algunos párrafos de un artículo titulado «The Emergence of Modern Neuroanatomy and Developmental Neurobiology» publicado por Cowan en el año 1998 en un número especial de la revista Neuron que lleva como título «One Decade of Neuron, Six Decades of Neuroscience» (3).
«... It is a measure of Cajal’s genius that he recognized that no single method could resolve this issue ...»
«... it is to Cajal and his brilliant use of the Golgi method that we owe our understanding of the development of individual neurons, the appearance of growth cones at the end of their processes, and most astonishingly, the microscopic appearance of virtually all regions in the developing brains of each of the major vertebrate classes... Cajal also put forward a number of remarkably prescient hypotheses concerning axon guidance, synapse formation, and the elimination of misplaced neurons and aberrant connections, as well as the latent capacity for regeneration and plasticity in the mature nervous system»
En un subcapítulo titulado «Axonal pathfinding and target recognition» dice: «... And, once again, it was Cajal who first clearly articulated the key questions: What causes the axon of a neuron to emerge from one or the other pole of the cell? How does the axon grow toward its target area through a myriad of other cells and process? How do axons that cross to the opposite side of the brain or spinal cord recognize and respond to cues at the midline and the contralateral side? How do axons identify their appropriate target cells, and how do they come to form synapses only upon the appropriate parts of those cells?»
Estas breves frases de Cowan resumen la grandeza y el porqué de la actualidad de la obra de Cajal. Sorprende que hace cien años Cajal se ocupara y orientara acertadamente sus investigaciones en estos temas de tan candente actualidad hoy, como si hubiese sido un científico que trabajara a principio de siglo XXI.
En los últimos veinte y cinco años he participado en numerosos actos, he dado conferencias, he organizado cursos y he escrito artículos en los que el tema de mi participación ha sido directa o indirectamente la actualidad de la obra científica de Cajal (4,5). En otras ocasiones me he limitado a advertir que ese concepto, ese experimento, esos resultados habían sido ya expuesto, planteado, obtenidos por Cajal, en algún caso cien años antes. Otras veces he señalado que Cajal había sido leído mal o incompletamente, pues precisamente Cajal decía todo lo contrario de lo que el conferenciante le atribuía; cosa que es frecuente en lo que a degeneración y regeneración en sistema nervioso central se refiere.
Es frecuente leer que se atribuye a Cajal el concepto de la incapacidad regenerativa del sistema nervioso central (4-7). Este es un ejemplo de que Cajal ha sido mal leído, o no ha sido leído, y apoyados en su autoridad se trasmiten frases supuestamente dichas por Cajal, copiándose unos autores a otros en el error, cuando él lo que defendió fue el concepto contrario: la capacidad intrínseca de regeneración de las neuronas del sistema nervioso central. Cajal en 1913 en su libro Estudios sobre la degeneración y regeneración en sistema nervioso (8) escribe en letra pequeña que las neuronas del sistema nervioso central tienen una incapacidad regenerativa. Sin embargo en la página anterior había afirmado en letra normal la opinión contraria y explica a continuación, también en letra normal, que no es una incapacidad intrínseca del sistema nervioso central, sino que son las condiciones fisicoquímicas que envuelven a sus neuronas las que condicionan esta situación, desapareciendo la misma cuando las circunstancias fisicoquímicas adversas desaparecen. Él en 1923 en Historia de mi labor científica (9), explica este tema con las siguientes frases:
«... la creación de retoños (trofismo) y su orientación al través de los diversos tejidos (tropismo) hállase condicionada por la liberación, en torno a las fibras y células, de fermentos activadores de la asimilación protoplásmica. Estos agentes catalíticos (substancias neurotrópicas) son fabricadas por el tejido conectivo embrionario; pero muy señaladamente por las células de Schwann...
En condiciones normales, los citados reclamos faltan... en cuanto concurren circunstancias experimentales favorables, la tendencia regenerativa, latente en las fibras de los centros, se despierta y alcanza extraordinaria pujanza.» (pág. 299).
«... Tales hechos, de gran trascendencia biológica, refutan definitivamente el dogma, generalmente admitido, de la irregenerabilidad esencial de las vías centrales. Tamaña incapacidad productiva constituye capacidad contingente y adventicia, motivada, según dejamos dicho, por la ausencia irremediable, dentro de la sustancia blanca y gris, de fuentes secretoras de agentes catalíticos y materias orientadoras*.»
«*... el crecimiento (trofismo) y orientación (tropismo) de los retoños nerviosos es función de las condiciones físico-químicas del medio. «(pág. 300).
En resumen para Cajal, la falta de procesos regenerativos en el Sistema Nervioso Central se debe a la ausencia de «fuentes secretoras de agentes catalíticos (capaces de crear retoños) –por tanto tróficos– y en materias orientadoras» (trópicas). Estos agentes tróficos y trópicos, según Cajal son fabricados por el tejido conectivo embrionario, pero muy señaladamente por las células de Schwamm en los tubos nerviosos en trance de regeneración. Si en el Sistema Nervioso Central se crean circunstancias experimentales favorables la tendencia latente se despierta. Esto lo mostraron Cajal y sus discípulos por medio de injertos y por medio de secciones simultáneas de cordones medulares y raíces sensitivas o motoras. Según Cajal, la degeneración de las células de Schwamm tanto en los injertos como en las secciones de raíces dan lugar a la liberación de sustancias que se difunden hasta las células del sistema nervioso central donde los axones cortados antes morosos e inertes crecen ahora activamente (función trófica) e invaden el injerto o las raíces en las que progresaron una larga distancia (función trópica) (fig. 2). Muy recientes hallazgos confirman la afirmación de Cajal de que trófico y trópico son dos lados de la misma moneda. Como afirma Víctor Hamburger (10) en 1980 en su magnífico ensayo sobre Ramón y Cajal «lo que fue visto por Cajal por pura reflexión, es decir que un agente promotor de crecimiento segregado por la estructura del destino, impregna los tejidos y guía la punta del crecimiento del axón (trópico) a su lugar de origen, ha sido ahora demostrado en un experimento controlado».

Fig. 2.
Son numerosos los hallazgos de los últimos quince años que demuestran las afirmaciones que Cajal hizo a principio de siglo sobre regeneración. Un ejemplo es un experimento en transplantado glía envolvente del bulbo olfatorio adulto (GE) a ambos lados de una sección medular y se han encontrado axones regenerados a larga distancia de la lesión con la misma localización que la GE (7). Concluyen sus autores que la GE proporciona a los axones lesionados con factores que permiten su elongación a larga distancia. Son en definitiva los agentes catalíticos y materias orientadoras, neurotróficos y neurotrópicas, de Cajal. Suponen que estas células gliales ofrecen nuevas posibilidades para el tratamiento de trastornos del sistema nervioso que requieran regeneración axonal. Estamos describiendo hechos que confirman los conceptos que expuso Cajal y que apenas han cambiado. Por ello no es extraño que Víctor Hamburger escribiera en 1991 (11): «Las ideas seminales de Cajal relativas a neurogénesis y regeneración nerviosa, que fueron concebidas alrededor del cambio de siglo, han servido de guía a tres generaciones de neurocientíficos, un testimonio duradero de su genio». Y yo en 1987 (5): ‘I think this article can best be concluded by recalling a Viktor Hamburger’s (10) sentence with an intercalated (underlined words) annotation: «… indeed, it is very difficult to be original in neurogenesis, regeneration and plasticity with Cajal looking over one’s shoulder».’ Hoy diría lo mismo.
La mayor parte de las descripciones de Cajal de la estructura microscópica del sistema nervioso central de los vertebrados siguen siendo de plena actualidad. Sus primeros trabajos con el método de Golgi sobre la estructura del cerebelo y la retina publicados en 1988 en la Revista trimestral de Histología normal y patológica y en la Gaceta Médica Catalana, en los que fundamentó la teoría neuronal, recogen todos los elementos nerviosos celulares y fibrilares que hoy son utilizados como esquemas por todas las revistas científicas, libros y artículos de revisión más autorizados, con los mismos nombres que dio Cajal. Naturalmente sin hacer referencia a Cajal. A título de ejemplo, entre miles, podemos examinar el esquema de la corteza cerebelosa en el trabajo de Mark Farrant en Nature (12) sobre receptores GABA y el esquema de la retina de la última edición del libro de Neurociencia de Kandel y colaboradores (13). Cajal editó en 1890 en Barcelona un opúsculo titulado «Textura de las circunvoluciones cerebrales de los mamíferos inferiores» en la que en imágenes de la corteza cerebral de ratones de pocos días, en dos grabados representaba en uno las neuronas de la capa I de la corteza cerebral y en el otro unas células gliales que desde la capa ventricular llegaban a la capa pial de la corteza y que él llamó células epiteliales (fig. 3). Las primeras neuronas son las que Retzius llamó neuronas de Cajal al describirlas un año más tarde en la corteza cerebral humana, y a las que Cajal definitivamente llamó células de Cajal-Retzius, nombre con el que han permanecido hasta ahora. Estas neuronas hasta hace diez años apenas habían sido motivo de especulaciones intranscendentes por algunos autores; sin embargo en los últimos años se ha demostrado que a través de la secreción de una glicoproteína, la reelina, son unos elementos esenciales en el desarrollo de la corteza cerebral, habiendo cobrado las células de Cajal-Retzius un protagonismo enorme en la literatura neurocientífica (14). Las células epiteliales de Cajal fueron bautizadas por Pasko Rakic (15) como glía radial, que sirve de guía a los neuroblastos que emigran desde la capa ventricular a la placa cortical cerebral, y cuya eficacia en esta función va a estar también condicionada por la reelina segregada por las células de Cajal-Retzius. Otros numerosos ejemplos podríamos poner en relación con este tema.

Fig. 3.
Opino que también resume bien el significado de la obra de Cajal una frase de mi prólogo a la obra de De Felipe y Jones «Cajal on the Cerebral Cortex» (4): Cajal’s posthumous work «¿Neurismo o Reticularismo?» can be considered his scientific testament. It offers the reader a unique panorama that helps us understand the persistence of Cajal’s work. Since 1888, when he defined the neuron doctrine, his scientific life was devoted to its defense. Toward his end, he carried out a genuine multidisciplinary effort. He studied almost every neural structure in different species, used a wide variety of the methods available in his time, and devised new experimental tools to pursue his goals».
Deseo insistir en una frase de este prólogo: «Toward his end, he carried out a genuine multidisciplinary effort.» Que Cowan repetiría diez años después de esta forma (3): «It is a measure of Cajal’s genius that he recognized that no single method could resolve this issue». Insisto en ello por que Cajal estudió el sistema nervioso con todas las técnicas que tuvo a su alcance y la Neurociencia actual se ocupa del estudio del sistema nervioso de una forma multidisciplinar. También en esto fue genial como recalca Max Cowan. Por ello no es extraño que muchos autores lo llamen el fundador de la moderna Neurociencia, algunos de los cuales hemos citado ya y expongo a continuación otros tres ejemplos: 1) Viktor Hamburger (10): «Ramón y Cajal es el fundador de la moderna neurología, que es también la base de la neurofisiología, neuropatología y psicología fisiológica.».... «su poder analítico intelectual al igual que su poder de observación. (....) Hay muy pocos problemas de la ciencia actual que no hayan sido tratados por él en una ocasión o en otra.».... «la recuperación de la idea de Cajal de que trófico y trópico deben de ser dos caras de la misma moneda puede ser un importante modelo en la neurobiología moderna del desarrollo. Verdaderamente, es muy difícil ser original en neurogénesis con Cajal mirando sobre nuestros hombros». 2) Jessell y Kandel en la Introduction al número de Neuron «One Decade of Neuron, Six Decades of Neuroscience» dicen (16): «the classical era of neural science, at the turn of the century, gave rise to the neuron doctrine and the concept of synaptic transmission by integrating information derived from the anatomical studies of Santiago Ramón y Cajal and the physiology of Charles Sherrington,». 3) Caroni (17): «The nervous system is made of billions of neurons that are interconnected with astonishing precision. How is genetic information decoded to wire this most complex of computing machines? By the end of the last century, the founder of modern neuroscience, Ramón y Cajal, had already proposed that the tips of the growing neuronal processes, which he called growth cones, made the navigational decisions required to specifically wire neuronal networks. Cajal also proposed that growth cones sampled their environment, guided in part by diffusible attractive factors».
Cajal modificó el concepto del sistema nervioso principalmente con la demostración y formulación de la teoría neuronal. Cajal descubrió la «nueva verdad» en el año 1888 con una técnica, el método de Golgi, que había aprendido un año antes. Hizo su presentación en sociedad el año 1989 en un congreso de la Sociedad Anatómica Alemana en la Universidad de Berlín. Es interesantísimo e instructivo leer el capítulo VI de «Historia de mi labor científica» (10) y no quiero dejar de transcribir los párrafos referentes a su demostración: «Desde muy temprano me instalé en la sala laboratorio —— enfoqué los cortes más expresivos. —— Entre los que más interés mostraron por mis demostraciones debo citar a His, Schwalhe, Retzius, Waldeyer y singularmente a Kölliker. —— iniciaron su examen con más escepticismo que curiosidad. —— Mas cuando hubieron desfilado ante sus ojos, en cortejo de imágenes clarísimas e irreprochables, el axón de los granos del cerebelo, las cestas pericelulares, las fibras musgosas y trepadoras, las bifurcaciones y las ramas ascendente y descente de las raices sensitivas, las colaterales largas y cortas de los cordones de la sustancia blanca, las terminaciones de las fibras retinianas en el lóbulo óptico, etc. —— las felicitaciones estallaron calurosas y sinceras». La Universidad de Berlín quiso celebrar el centenario de la puesta de largo de la teoría neuronal en aquella ciudad, que debido a las circunstancias especiales de Berlín hubo de retrasarse un año. Fui invitado a participar en esta celebración, lo que hice con una conferencia titulada «Cajal’s work after a century. Insightful ideas of a scientific genius», que pronuncié en la Virchow Hörsaal del Anatomischen Institut der Humboldt Universität el mes de octubre del año 1990. Mi emoción fue enorme al rememorar las vivencias de D. Santiago en aquellas salas y laboratorios que ese día rendían al sabio español un reconocimiento internacional.
Es la teoría neuronal base de todos nuestros conocimientos sobre el sistema nervioso, su morfología, su fisiología y su patología. Es base de su bioquímica. Y precisamente la individualidad anatómica y funcional de cada neurona, incluso con la que tiene más próxima, sus conexiones distintas, es lo que hace el estudio del sistema nervioso tan complejo y sólo hoy día con técnicas muy sofisticadas nos estamos aproximando a la caracterización morfológico-funcional y bioquímica de cada neurona o grupo de neuronas.
¿Cómo pudo llegar Cajal a la formulación de la teoría neuronal? Según él mismo señala con la observación de las estructuras nerviosas en estadíos más sencillos, es decir, el estudio del sistema nervioso en periodos embrionarios y en animales jóvenes. Además modificó el método de Golgi, introduciendo la doble impregnación.
En relación con la ontogenia el problema más urgente a resolver era averiguar cómo se forman los nervios y en virtud de qué mecanismos los apéndices axónicos se enlazaban «sin errores ni extravíos» con sus aparatos terminales. En aquel momento, años 1890, dos teorías explicaban este fenómeno, las de Hensen y la de Beard, Dohrn y Bethe. Cajal demostró de forma inconmovible la unidad genética de las fibras nerviosas y los apéndices protoplasmáticos, haciendo observaciones en embriones de pollo desde el segundo día de la incubación teñidas con el método de Golgi. Demostró que el neuroblasto emite primeramente el axón y sólo más tarde produce las prolongaciones protoplasmáticas y colaterales. «Contempló» y describió por primera vez el «cono de crecimiento». Comparable a un «ariete vivo», blando y maleable que avanza empujando mecánicamente los obstáculos hallados en su camino hasta asaltar su distrito de terminación periférica». Conos de crecimiento que volvería a estudiar en 1906 y 1908 con el método de nitrato de plata reducido, cuando de nuevo tuvo que defender la teoría neuronal, ante los ataques de Held y Apathy.
También en 1890 (18) describió Cajal la evolución ontogenética de las células y fibras de la corteza cerebelosa. Además de haber descrito el desarrollo de los granos del cerebelo, Cajal estudió el desarrollo de las células de Purkinje y cómo llegan a esas neuronas y trepan por ellas las fibras trepadoras. En el primer caso describió un fenómeno que hoy ha adquirido enorme importancia en el estudio de la plasticidad del sistema nervioso y que explica en gran parte la multipotencialidad de algunas neuronas en periodos jóvenes para fijarse su función en estados adultos. Explicó como en una primera fase surge un ramaje dendrítico abundante y multidireccional, que luego va a quedar reducido y orientado de acuerdo con las aferentes útiles al destino funcional de la neurona que crecen y se desarrollan despareciendo las prolongaciones inútiles, «como el jardinero poda las ramas superfluas». También señaló cómo los axones, hoy sabemos que son procedentes de las neuronas de la oliva inferior, según Cajal «olfatean el soma de los elementos de Purkinje, al cual abrazan mediante nidos varicosos ... las ramas del nido trepan positivamente a lo largo del tallo principal y dendritas (de las células de Purkinje) hasta generar el plexo complicado característico de los conductores adultos».
Cajal supo darle vida a las observaciones fijadas en las preparaciones histológicas; son palabras suyas: olfatean y trepan. Estas palabras recuerdan a la que hemos señalado más arriba al referirse al avance del «cono de crecimiento». O en las que se refiere en otra ocasión a esta misma estructura en 1906. El dinamismo de estas descripciones nos hace contemplar a los axones en crecimiento, pero igualmente nos llevan de la mano hacia otra de sus geniales propuestas: la teoría del neurotropismo. Las frases: «olfatean el soma de las células de Purkinje» y «con exquisita sensibilidad química» están hablando del problema de cómo los nervios encuentran su camino hacia su destino. Las primeras sugerencias de Cajal en este sentido se hicieron a propósito de la emigración de los granos del cerebelo embrionario de las capas superficiales a las más profundas. Sin embargo, como he señalado más arriba, es con el estudio de la regeneración de las fibras nerviosas cuando Cajal elabora en toda su extensión esta teoría. Cajal estudió la regeneración de los nervios en el Sistema Nervioso Periférico y Sistema Nervioso Central, utilizando su nueva técnica de nitrato reducido durante los años 1905 y 1906 con la que también estudió estructuras del sistema nervioso central, en algunas de las cuales son sus imágenes y descripciones las únicas referencia que encuentra el investigador que pretende estudiarla a nivel de microscopía electrónica (fig. 4). Todos estos hallazgos justifican algunas de las afirmaciones y preguntas que Cowan (3) atribuía a Cajal y las referencias de Viktor Hamburger (10), Jessell y Kandel (16) y Caroni (17) que he trascrito en páginas anteriores.

Fig. 4.
Para terminar con este capítulo de neurogénesis desearía hacer un comentario a la pregunta de Cajal formulada más arriba por Cowan (3) de ¿Que hace el axón para cruzar al otro lado del cerebro o médula espinal reconociendo y respondiendo a señales de la línea media y del lado contralateral? Hay numerosas imágenes de Cajal sobre las que planteó esta pregunta que pueden compararse a algunas de las que muestran la bibliografía más actual, por ejemplo con axones de células derivadas de la placa alar de la médula espinal atravesando la línea media por la placa del suelo y otros quedándose en el mismo lado (fig. 5) (19). Hoy conocemos algunas de esas señales que preconizaba Cajal. Sabemos que la placa del suelo segrega una sustancia atrayente, netrina, y una repelente Robo (20). Los axones que expresan un alto nivel de Robo no cruzan la línea media y los que la han cruzado expresarán Robo después de cruzarla para quedarse en ese lado.
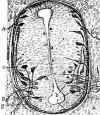
Fig. 5.
Finalmente, haré algún comentario sobre otro hallazgo de Cajal que descubre una vez más su capacidad de plantear experimentos originales y de interpretar los hechos de forma sencilla, permanente. Me refiero a sus estudios de las reacciones, él llama «adaptación morfológica de las neuronas a las condiciones fisiológicas artificiales provocadas por la mutilación» que es una clara demostración de la plasticidad del sistema nervioso central. Cajal concluye con una frase que hoy nos suena a total actualidad: «la morfología de las células nerviosas no obedece a causa inminente y fatal, mantenidas por herencia sino que depende enteramente de las circunstancias actuales físicas y químicas del ambiente» (9). Hoy decimos genética y ambiente condicionan el desarrollo y comportamiento de una estructura nerviosa (21). Sólo un ejemplo de sus extensos trabajos en este sentido publicados en 1911 (22): a una célula de Purkinje Cajal cortó sus axones distalmente a la salida de colaterales (fig. 6). Como consecuencia sucedieron dos fenómenos: 1.º Estas células dejaron de proyectar fuera de la corteza cerebelosa. 2.º Las colaterales que quedaron se hipertrofiaron y parece han ampliado su área de terminación. Cajal comenta el primer fenómeno y dice que hemos transformado una célula de axón largo en una célula de axón corto. Las posibles previsiones genéticas han quedado modificadas por las circunstancias ambientales que se han dado en este caso en el cerebelo (8). El segundo fenómeno, el mayor desarrollo de las colaterales podría explicarse al ser ellas ahora las únicas prolongaciones que dependen de la actividad trófica de las neuronas es decir, estamos ante un «pruning effect», descrito en el último tercio de siglo XX (23). Además, las colaterales ocupan ahora los espacios sinápticos vacíos que han dejado las fibras trepadoras cortadas al seccionar los axones de las células de Purkinje. Fenómeno también redescubierto a finales del pasado siglo. Descripciones semejantes hizo en la corteza cerebral.

Fig. 6.
Pero no quería dejar de insistir en la afirmación que copiaba de Viktor Hamburger unos párrafos anteriores: «Hay muy pocos problemas en la Neurociencia actual que no hayan sido tratados por Cajal en una ocasión u otra». Solamente citaré dos ejemplos. El primero es una frase que los tres autores más autorizados en el estudio de los mecanismos del aprendizaje y la memoria a nivel básico y clínico escribían a propósito de «Aproximación Molecular y Biológica al Depósito de la Memoria» (24): «Este hecho animó a Santiago Ramón y Cajal (1894) a proponer que el aprendizaje ... hace que células nerviosas existentes emitan o hagan crecer nuevas prolongaciones para reforzar sus conexiones con otras células nerviosas, así que sea posible comunicarse con ellas más eficazmente». La parte fisiológica de esta afirmación había hasta ahora podido ser demostrada en invertebrados y mamíferos, buen ejemplo de lo segundo es la potenciación a largo plazo que ha sido demostrada en el hipocampo y otras regiones del sistema nervioso central de mamíferos (25). Pero el aspecto morfológico-estructural de esta afirmación de Cajal solamente había podido ser demostrado en invertebrados; sin embargo, recientemente lo ha sido también en mamíferos (26). El segundo como una demostración de la amplitud en los campos de la neurociencia que ha influido Cajal citaré una frase de un artículo de Nicoll en Nature titulado «Cajal’s rational psychology» (27): ‘... the brain’s intricate cellular architecture is best summarized by the most famous of neuroanatomists, Ramón y Cajal: «... We saw that an exact knowledge of the structure of the brain was of supreme interest for the building up of a rational psychology.» ...Perhaps we are not, after all, that far away from Cajal’s rational psychology.’
Muchos hechos objetivos, como vemos, demuestran efectivamente que la obra de Cajal sigue siendo actual y uno singularmente demostrativo es que Cajal es uno de los científicos más citados en Neurociencia. Las citas de sus trabajos, lejos de disminuir, aumentan en los últimos años (próximas a 500 anuales). Esto es más asombroso cuando hoy la mayoría de los trabajos científicos quedan obsoletos en tres o cuatro años. Estamos pues no sólo ante una obra genial, sino ante unas descripciones tan sólidamente fundadas que más de cien años después permanecen y estoy seguro permanecerán siempre.
Para analizar el porqué de la permanencia de la obra de Cajal debemos tener en cuenta que era un hombre genial, que tenía un gran espíritu crítico y una enorme capacidad para interpretar las imágenes observadas en el microscopio. Sin embargo, el secreto de la permanencia de la obra cajaliana está en esos conceptos que él repite en muchas ocasiones con palabras diferentes (9): «los hechos quedan, las teorías pasan», «la fama duradera sólo acompaña a la verdad», etc. Es decir fidelidad en representar y describir lo que se ha visto. Esta fiabilidad de la obra de Cajal es la principal causa de su permanencia. Sin embargo, se necesitaron unir a ella las cualidades de capacidad intelectual y su gran habilidad y capacidad para representar gráficamente y con enorme fidelidad lo que observaba e el microscopio, del que, como él mismo describe, era «un obrero infatigable». Si todas las cualidades más arriba mencionadas hacen de su obra permanente, está última, su infatigabilidad en mirar por el microscopio, su enorme capacidad de trabajo, su entusiasmo y pasión por la investigación neuroanatómica hacen además su obra enorme, extensa. Así fue una obra extensa y permanente que inauguraba toda una etapa del conocimiento del sistema nervioso en la que seguimos haciendo la referencia a Cajal imprescindible en la mayor parte de los temas que se aborden en la investigación neurobiológica. Sus «conquistas científicas creaciones de su voluntad y ofrendas de su pasión» siguen siendo punto de partida de gran parte de nuestros trabajos.
Los hallazgos de Cajal y sus interpretaciones nos introducen aún en el terreno conceptual en múltiples campos que han comenzado su desarrollo en épocas muy recientes. Por eso no nos extraña que Cajal siga siendo un autor muy citado, no sólo cuando se trata de temas de morfología y conectividad del Sistema Nervioso, sino también cuando se trata de técnicas de tinción, embriogénesis, desarrollo, plasticidad, transplantes y otros muchos actuales.
BIBLIOGRAFÍA
F. Reinoso-Suárez. Introducción. En: Recuerdos de mi vida: Historia de mi labor científica. S. Ramón y Cajal. Alianza Editorial, Madrid, pp. 17-23, 1981.
F. Reinoso-Suárez et al., Eu J Neurosci 6:1829-1836, 1994
Neuron 20: 413-426
Algunos ejemplos: F. Reinoso-Suárez. Ramón y Cajal’s
Contribution to the Neuroscience. En Cajal: A modern insight in
neuroscience. Eds. S. Grisolía, Guerri, Samson, Norton, F. Reinoso-Suárez.
Elsevier Science Publishers, B.V., pp. 3-22, 1983.
F. Reinoso-Suárez. Actualidad de la obra científica de Cajal. Arbor, 114:
305-313, 1983.
F. Reinoso-Suárez. Introduction. En: Cortical Integration: Basic,
archicortical and cortical association levels of neural integration. Eds. F.
Reinoso-Suárez, C. Ajmone Marsan. IBRO Monograph Series, Raven Press, New
York, vol. 11, pp. 1-11, 1984.
F. Reinoso-Suárez. Foreword. En: Cajal on the Cerebral Cortex. J. DeFelipe
and E.G. Jones. Oxford University Press, New York and Oxford, pp. VII-IX,
1988.
F. Reinoso-Suárez. Actualidad de la obra científica de Cajal. Madrid
Médico, 114: 42-45, 1989.
F. Reinoso-Suárez. Cajal y la Neurociencia en el umbral del tercer milenio.
An R Acad Nac Med (Madr) 117: 451-467, 2000.
F. Reinoso-Suárez. Cajal’s concept on plasticity in the central nervous system revisited: a perspective. En: Neuroplasticity: new therapeutic tool in the CNS pathology. Eds. R.L. Masland, A. Portera-Sánchez, G. Toffano. Fidia research series, vol. 12, Liviana Press. Padova, pp. 31-37, 1987.
F. E. Bloom. CNS Plasticity: A Surey of Opportunities. En: Central Nervous System Plasticity and Repair. Eds: A. Bibnami, C. L. Bolis, F. E. Bloom and A. Adeloye. Raven Press New York, pp: 3-11, 1985.
A. Ramón-Cueto, G. W. Plant, J. Avila, M. B. Bunge. Long-distance axonal regeneration in the transected adult rat spinal cord is promoted by olfactory ensheathing glia transpants. J. Neurosci. 18: 3803-3815, 1998.
S. R. y Cajal. Estudios sobre la degeneración y regeneración en sistema nervioso. N. Moya, Madrid, 1913-1914.
S. R. y Cajal. Recuerdos de mi vida: Historia de mi labor científica. Reeditada por Alianza Editorial, Madrid, 1981.
V. Hamburger. S. Ramón y Cajal, R. W. Harrison and the beginnings of neuroembriology. Persp. Biol. Med. 23: 600-616, 1980.
V. Hamburger, Foreword. En: Cajal’s Degeneration and Regeneration of the Nervous System, 1991.
M. Farrant, S. Cul-Candy, Neurobiology: GABA receptors, granule cells and genes. Nature, 361: 302-303, 1993.
M Tessier-Lavigne, Visual processing by the retina. En: Principles of Neural Science. Eds. E.R. Kandel, J.H. Schartz, T.M. Jessell. McGraw-Hill, New York, p. 515, 2000.
A.M. Goffinet, Unscrambling a disabled brain. Nature, 389: 668-669, 1997.
P. Rakic, Mode of cell migration to the superficial layers of fetal monkey neocortex. J. Comp. Neurol., 146: 335-345, 1972.
Neuron 20:367-369, 1998
Caroni, P., Driving the growth cone. Science, 281: 1465-1466, 1998.
S. R. y Cajal. Sur les fibres nerveuses de la couche granuleuse du cervelet et sur l’évolution des éléments cérébelleux. Monatschr. Anat. Physiol. 1890. Reimpresión en Trabajos Escogidos (1924) Vol. 1. Jiménez y Molina, Madrid, pp. 451-466.
S. R. y Cajal. Génesis de las fibras nerviosas del embrión y observaciones contrarias a la teoría catenaria. Trab. Lab. Invest. Biol. 4: 219-284, 1906.
B. Dickson, A roundabout way of avoiding the midline. Nature, 391: 442-443, 1998.
E.R. Kandel, J.H. Schartz, T.M. Jessell. Principles of Neural Science. McGraw-Hill, New York, 2000.
S. R. y Cajal, Reacciones degenerativas de las células de Purkinje del cerebelo bajo la acción del traumatismo. Bol.Soc. Esp. Biol. Sesión del 21 de abril de 1911.
G. E. Schneider, Early lesion of superior colliculus: factors affecting the formation of abnormal retinal projections. Brain Behav. Evol. 8: 73-109, 1973.
B. Milner, L.R. Squire and E.R. Kandel, Cognitive Neuroscience and the Study of Memory. Neuron, 20: 445-468, 1998.
T.V.P. Bliss, G.L. Collingridge, A synaptic model of memory: Long-term potentiation in the hippocampus. Nature, 361: 31-39, 1993.
M. Barinaga, Learning visualized, on the double. Science , 286: 1661, 1999.
R.A. Nicoll, Nature 368: 808-809, 1994.
![]()