
Vol. 35, n.º 4, 2002
|
REVISTA
ESPAÑOLA DE
Vol. 35, n.º 4, 2002 |
Santiago Ramón y Cajal
Mesdames, Messieurs,
Conformément à la tradition à laquelle sont restés fidèles les illustres conférenciers honorés avant moi du prix Nobel, je vais vous entretenir des principaux résultats de mes travaux scientifiques dans les domaines de l’histologie et de la physiologie du système nerveux.
De l’ensemble de mes recherches découle une conception générale qui comprend les propositions suivantes:
I. Les cellules nerveuses sont des individualités morphologiques, des neurones, suivant le mot consacré par l’autorité du prof. Waldeyer. Cette propriété fut déjà démontrée par mon illustre confrère le prof. Golgi à l’égard des prolongements dendritiques ou protoplasmiques des cellules nerveuses; mais en ce qui concerne la façon de se comporter des dernières ramilles des axons et des collatérales nerveuses, il n’y avait, au début de nos recherches, que des conjectures plus ou moins soutenables. Nos observations avec la méthode de Golgi, que nous appliquâmes, d’abord dans le cervelet, ensuite dans la moelle épinière, le cerveau, le bulbe olfactif, le lobe optique, la rétine etc des em bryons et des jeunes animaux, révélèrent, à mon avis, la disposition terminale des fibres nerveuses. Celles-ci, en se ramifiant à plusieurs reprises, se portent constamment vers le corps neuronal ou vers les expansions protoplasmiques autour desquels prennent naissance des plexus ou des nids nerveux très serrés et fort riches. Les corbeilles péricellulaires et les plexus grimpants et d’autres dispositions morphologiques, dont la forme varie selon les centres nerveux que l’on étudie, attestent que les éléments nerveux possèdent des relations réciproques de contiguité et non de continuité, et que ces rapports de contact plus ou moins intime s’établissent toujours, non entre les arborisations nerveuses seules, mais entre ces ramifications d’une part et le corps et les prolongements protoplasmiques d’autre part. Un ciment granuleux ou substance conductrice particulière servirait à relier très intimement les surfaces neuronales en contact.
Ces faits reconnus dans tous les centres nerveux à l’aide de deux méthodes très différentes (celle de Golgi et celle D'Ehrlich), confirmés et notablement développés par les recherches de Koelliker, v. Lenhossék, Retzius, van Gehuchten, Lugaro, Held, mon frère, Athias, Edinger, et bien d’autres, impliquent trois postulats physiologiques:
1) Puisque la nature, afin d’assurer et d’amplifier les contacts, a créé des systèmes compliqués de ramifications péricellulaires (systèmes qui deviendraient incompréhensibles dans l’hypothèse de la continuité), il faut admettre que les courants nerveux se transmettent d’un élément à l’autre en vertu d’une sorte d’induction ou d’influence à distance.
2) Il faut aussi supposer que les corps cellulaires et les prolongements dendritiques sont, de même que les cylindres axes, des appareils de conduction, puisqu’ils représentent des anneaux intermédiaires entre les fibres nerveuses afférentes et les axons mentionnés. C’est ce que Bethe, Simarro, Donaggio, nous-même, etc., avons confirmé, tout récemment, en démontrant, à l'aide des méthodes neurofibrillaires, une parfaite concordance structurale entre les dendrites et le prolongement cylindre-axile.
3) L’examen de la marche des impulsions nerveuses dans les organes sensoriels, tels que la rétine, le bulbe olfactif, les ganglions sensitifs et la moelle épinière, etc., prouve, non seulement que les expansions protoplasmiques jouent un rôle conducteur, mais encore que le mouvement nerveux est dans ces prolongements cellulipète ou axipète, tandis qu'il est cellulifuge dans les axons. Cette formule, dite de la polarisation dynamique des neurones, créée il y a longtemps par van Gehuchten et nous comme induction tirée de nombreux faits morphologiques, n’est pas en contradiction avec les nouvelles recherches sur la constitution du protoplasma nerveux. Nous verrons en effet que la charpente neurofibrillaire constitue un réticulum continu depuis les dendrites et le corps cellulaire jusqu'à l'axon et sa terminaison périphérique.
Pendant vingt-cinq années de labeur continu, portant sur presque tous les organes du système nerveux et sur un grand nombre d’espèces zoologiques je n’ai jamais rencontré un seul fait d’observation contraire à ces assertions, bien que j’aie employé dans mes recherches, outre les procédés communs de coloration, les méthodes électives de Golgi, Cox, Ehrlich et tout dernièrement les méthodes neurofibrillaires. Ajoutons que la même doctrine découle aussi de l’ensemble des observations de Kœlliker, v. Lenhossék, van Gehuchten, mon frère, Edinger, Lugaro, etc, sur le système nerveux des vertébrés, et de celles très importantes de Retzius sur le système nerveux des invertébrés.
Je devrais maintenant passer en revue les principaux faits d’observation sur lesquels les précédentes inductions sont basées. Ce serait aussi le meilleur moyen de vous montrer la portée et l’étendue de ma modeste contribution personnelle à la connaissance du système nerveux. Malheureusement il est absolument impossible de résumer en quelques pages des faits morphologiques dont la description occupe un grand nombre de brochures avec des centaines de dessins. Cependant, afin de ne pas fatiguer outre mesure votre très bienveillante attention, je me bornerai à choisir dans l’ensemble de mes travaux quelques exemples frappants de connexion interneuronale que j’ai reproduits schématiquement dans les tableaux que voici:
Voyons tout d’abord les connexions des racines sensitives de la moelle épinière. On sait bien, d’après les recherches de Ranvier, de Retzius, et de v. Lenhossék, etc. que le prolongement unique des corpuscules sensitifs est divisé en deux branches: l'une externe qui se dirige vers la périphérie pour se terminer dans la peau ou dans les muqueuses; l’autre interne qui pénètre dans la racine sensitive ou postérieure pour se rendre au cordon dorsal de la moelle épinière. Cette dernière branche, d’après mes observations chez les oiseaux, chez les reptiles et chez les mammifères (confirmées par un grand nombre de savants tels que Kœlliker, v. Lenhossék, Retzius, van Gehuchten, Sala, Athias, etc.), ne pénètre pas directement, ainsi que l’avaient supposé certains auteurs, dans la substance grise, mais se divise dans l’épaisseur du cordon postérieur de façon à donner une branche ascendante et une autre descendante (fig. 1A). Cette bifurcation a la forme d’un Y et les fibres qui en résultent se portent le long du cordon dorsal pendant un trajet considérable, pour se terminer, en définitive, dans le sein de la substance grise au moyen d’arborisations variqueuses et péricellulaires.
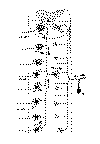
Fig. 1.
Mais outre ces arborisations terminales, les fibres radiculaires sensitives offrent aussi un nombre considérable de ramilles collatérales partant à angle droit, soit de la tige, soit des branches ascendantes et descendantes, grâce auxquelles elles se mettent en connexion avec tous les neurones de la substance grise (fig. 1C, 2 a, b et c). On peut distinguer ces collatérales en deux variétés principales: de longues ou réflexo-motrices, destinées à se mettre en contact avec les neurones moteurs (fig. 2c) ainsi que nos recherches et surtout celles de Kœlliker et de v. Lenhossék l’ont bien démontré; et de courtes, destinées à se mettre en relation avec les neurones funiculaires (homolatérales et opposito-latérales) qui siègent dans les deux cornes de la substance grise (fig. 2 a et c).
Les arborisations terminales de ces fibres produisent autour des neurones et de leurs dendrites des plexus terminaux très serrés, apparents surtout au niveau des corpuscules moteurs. Nos recherches, portant d’abord sur les em bryons et les animaux àgés de quelques jours, avaient signalé dans ces nids des varicosités terminales; tout dernièrement Held et Auerbach dont les recherches ont été confirmées et complétées par nous, van Gehuchten, Mahaim, Holmgren et bien d'autres à l'aide de notre méthode de l’argent réduit, ont démontré que ces varicosités sont notablement développées chez les mammifères adultes et contiennent un réseau ou un anneau neurofibrillaire (1).
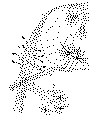
Fig. 2.
Comme le montre la figure 2 c, le mouvement nerveux amené par les radiculaires sensitives, se divise en deux courants importants: le courant direct ou réflexo-moteur direct qui se propage sans neurone intermédiaire dès la racine postérieure jusqu’aux neurones moteurs; et le courant réflexo-moteur indirect ou associatif, lequel se rend aux neurones moteurs très éloignés, mais après avoir fait un détour en passant par un corpuscule nerveux intermédiaire ou d’association, c’est-à-dire par les neurones funiculaires directs ou commissurels dont l’axon, d’après nos observations, se divise très souvent dans la substance blanche, en produisant une branche ascendante et une autre descendante (fig. 1 C).
Dans la figure 3 je montre les connexions des fibres visuelles et des cellules de la rétine, objet d’étude où les rapports interneuronaux se présentent avec une admirable simplicité et netteté. Malgré sa grande complication, la rétine peut être considérée comme un ganglion nerveux formé par trois rangées de neurones ou de corpuscules nerveux: la première rangée renferme les cônes et les bâtonnets avec leurs prolongements descendants produisant la couche des grains externes (fig. 3 a, b), la seconde est constituée par les cellules bipolaires (c, d), et la troisième contient les neurones ganglionnaires (e); les trois séries de corpuscules nerveux s’articulent au niveau des couches dites moléculaires ou plexiformes externe et interne.
Notons que la conche plexiforme externe (fig. 3 C) renferme une articulation multiple dont les éléments sont: en dehors, les sphérules terminales de la fibre des bâtonnets et les pieds coniques munis d’excroissances filamenteuses des prolongements descendants des cônes; en dedans, les panaches externes des cellules bipolaires, dont, comme nous avons signalé, il existe deux variétés: les bipolaires à panache aplati ou destinées aux cônes (fig. 3 d), les bipolaires robustes à panache dendritique ascendant ou destinées aux bâtonnets (fig. 3 c), et enfin les branches protoplasmiques et les arborisations nerveuses des cellules horizontales de la couche des grains internes.
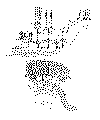
Fig. 3.
La couche plexiforme interne offre une articulation encore plus compliquée que l’on pourrait décomposer en trois ou en un plus grand nombre d’étages. Les facteurs essentiels sont representés: en dehors, par les panaches terminaux du prolongement descendant des bipolaires et les ramifications terminales de l’expansion inférieure des spongioblastes; en dedans par les arborisations protoplasmiques aplaties des neurones de la zone ganglionnaire.
En poursuivant les axons des neurones de la couche ganglionnaire au long du nerf optique, nous trouverons encore dans le cerveau moyen et le cerveau intermédiaire une troisième articulation mise en lumière d’abord par nos recherches sur le lobe optique des oiseaux et le cer veau moyen des mammifères, puis par les très intéressantes observations de mon frère (corps genouillé externe des mammifères, lobe optique etc des oiseaux, des reptiles et des poissons) et celles de van Gehuchten, Kœlliker, Sala, Tello etc. Ainsi que l'on sait, certains cylindres axes de la bandelette optique se portent en avant se terminant par des ramifications libres très compliquées, dans l’épaisseur du corps genouillé externe, d’autres, se dirigeant en arrière, se décomposent en de magnifiques arborisations ascendantes dans l’écorce du tubercule quadrijumeau antérieur, où se trouve l’origine de la voie visuelle réflexo-motrice (fig. 3 g).
Enfin des observations effectuées chez les mammifères et chez l’enfant nous ont démontré la quatrième et dernière articulation des conducteurs optiques, c’est-à-dire de la voie optique centrale dont les neurones d’origine siègent dans le corps genouillé externe. Cette intéressante connexion terminale, contrôlée par les importants travaux anatomo-pathologiques de Henschen, a lieu dans la scissure calcarine au niveau de la 4e et 5e zone corticale dans laquelle se trouvent deux couches très serrées de cellules étoilées (fig. 7 g).

Fig. 4.
Voyons maintenant dans les figures 4 et 5 quels sont les neurones et les connexions des cellules et des fibres dans une lamelle cérébel leuse. Comme on sait bien, une coupe transversale des ces lamelles nous montre trois couches concentriques de neurones.
La première ou zone plexiforme est formée principalement par les petites cellules étoilées (cellules à corbeilles de quelques auteurs). La seconde ou intermédiaire est constituée par les corps des cellules de Purkinje. La troisième, enfin, résulte de la réunion des grains.
Il ressort de mes observations confirmées par un grand nombre d’autours (van Gehuchten, Kœlliker, Lugaro, Edinger, mon frère, Falcone, Retzius, Azoulay, Held, etc.) que tous ces éléments possèdent deux espèces de rapports: des connexions intrinsèques, c’est-à-dire établies entre les neurones siégeant dans des zones superposées, et des relations extrinsèques, ayant lieu entre les neurones du cervelet et des cellules nerveuses appartenant à d’autres organes centraux.
Examinons d’abord les connexions intrinsèques et commençons par celles ayant lieu entre les corpuscules de Purkinje et les axons des petits éléments étoilés de la couche plexiforme.
Ces axons émettent plusieurs collatérales et, après un parcours variable, décrivent une courbe pour venir s’achever au niveau des corps des neurones de Purkinje, à l’aide d’un grand nombre de ramifications successivement épaissies (fig. 4 A). Ces branches terminales, de même que les ramilles issues des collatérales descendantes, constituent autour des somas de Purkinje, un nid ou plexus serré qui souvent se termine inférieurement en pointe de pinceau (fig. 4B). L’intéressante connexion par contact qui s’établit de cette façon entre ces deux ordres de neurones a été confirmée à l’aide du bleu de méthylène par Dogiel et par nous. Tout dernièrement nous-même, Bielschowski, Wolff, et bien d’autres avons réussi à la démontrer admirablement imprégnée avec les méthodes neurofibrillaires.
Voyons à présent les rapports entre les grains et les cellules de Purkinje. Les grains sont de petits éléments nerveux, munis de quelques fines dendrites terminées par une ramification digitiforme, et d’un axon extraordinairement délicat (fig. 5 a). Ce prolongement nerveux monte jusqu’à la zone plexiforme et en s’y bifurquant à différentes hauteurs produit une fibre très délicate dite parallèle, parce qu’elle est disposée en une direction parallèle à celle des lamelles du cervelet. Pendant son long trajet longitudinal cette fibre se met en contact avec les contours épineux des branches des dendrites des cellules de Purkinje. Comme chaque fibrille parallèle parcourt la longueur totale d’une lamelle cérébelleuse, il s’ensuit qu’un seul grain peut agir sur une multitude de cellules de Purkinje.

Fig. 5.
Ajoutons encore parmi les rapports intrinsèques ceux établis entre les collatérales récurrentes des corpuscules de Purkinje et les grosses branches dendritiques de ces derniers (fig. 4 b). Ainsi que l’on sait bien, les collatérales récurrentes découvertes par Golgi se portent à la couche moléculaire dans laquelle elles se ramifient à plusieurs reprises. Pendant longtemps on a ignoré la façon de se terminer de ces branches nerveuses, et on avait imaginé bien des suppositions concernant leurs connexions avec les éléments de la couche plexiforme. Tout dernièrement en étudiant le cervelet du chien et de l’homme à l’aide du nitrate d’argent réduit, nous avons eu la chance de découvrir que les dernières ramilles des dites fibres, après être devenues longitudinales, se terminent au moyen d’un minuscule anneau neurofibrillaire sur la surface des tiges dendritiques des corpuscules de Purkinje (fig. 4 b). Ce fait montre donc, ainsi que nous l’avions admis jadis, que les collatérales récurrentes jonent le rôle d’associer en un ensemble dynamique les neurones congénères d’un même foyer de la substance grise (fig. 4 C).
Les rapports extrinsèques des cellules de l’écorce cérébelleuse ne sont connus qu’imparfaitement. Nous savons, ainsi que Golgi le démontra, que les cellules de Purkinje donnent naissance à des prolongements nerveux du type long ou moteur dont la terminaison est ignorée (probablement elle se trouve dans l’olive cérébelleuse et le ganglion du toit). Et inversement nous connaissons la façon de se terminer dans le cervelet de deux espèces de fibres nerveuses afférentes, les fibres moussues et les fibres grimpantes dont les neurones d’origine sont encore énigmatiques.
D’ailleurs, les connexions établies entre ces deux classes de conducteurs et les cellules de l’écorce cérébelleuse, sont très intéressantes pour la théorie. Elles ont contribué beaucoup à nous persuader de la légitimité de la doctrine neuronale; d’autant plus que les images de ces rapports, révélées d’abord par la méthode de GOLGI, nous les avons confirmées pleinement en nous servant des procédés d’Ehrlich et du nitrate d’argent réduit.
Vous savez bien que les fibres moussues sont de gros tubes médullaires qui se ramifient et se terminent dans la couche des grains en se mettant en contact, à l’aide de leurs rosacés ou ramifications épaisses et variqueuses, avec les branches digitiformes des grains (fig. 5 A). Cette très curieuse articulation, signalée d’abord par nous (1894) et confirmée par Held, Berliner, Wolff etc., a lieu dans certains territoires d’aspect granuleux, dépourvus de noyaux, qu’on a appelés glomérules cérébelleux à cause de leur ressemblance avec les glomérules olfactifs. D’après nos récentes observations, confirmées par Bielschowsky et Wolff qui ont travaillé avec la méthode de ce premier auteur, les excroissances finales des fibres moussues présentent un réticulum neurofibrillaire lâche et même des anses et des anneaux terminaux.
Quant aux fibres grimpantes, elles traversent la couche des grains, longent les corps des cellules de Purkinje et enveloppent les tiges ascendantes et les branches secondaires principales de ces neurones d’une magnifique arborisation terminale allongée grimpante que l’on pourrait comparer à celle des fibres motrices sur les corpuscules musculaires striés (figs. 4 D et 5 C).
Il résulte de ce que nous venons d’exposer que les grains et les neurones de Purkinje peuvent recevoir des actions nerveuses d’autres centres, probablement des ganglions de la protubérance et de la branche ascendante du nerf vertibulaire; tandis que les gros éléments étoilés de la zone des grains semblent ne posséder aucune relation avec les fibres extrinsèques.
Nous n’avons pas le temps de passer en revue tous les exemples très convaincants d’articulation neuronale que nous offrent d’autres centres nerveux, tels que le bulbe olfactif, l’écorce cérébrale, le thalame optique, les ganglions sensitifs et sympathiques, etc., sujets d’étude sur lesquels nous avons fait jadis un grand nombre de recherches. Nous nous bornerons ici à mentionner succinctement l’existence d’un facteur spécial des articulations intercellulaires et dont le rôle physiologique, encore fort discuté, doit avoir une grande importance. Nous faisons allusion aux fibres centrifuges, signalées il y a longtemps par nous et par Dogiel dans la rétine, puis retrouvées dans le bulbe olfactif, le tubercule quadrijumeau antérieur et le thalame optique, dans lequel elles sont particulièrement abondantes.
Ainsi que l’on peut remarquer dans la figure 6, a, qui représente une coupe schématique de la rétine des oiseaux, des fibres centrifuges, provenant de centres nerveux encore inconnus, traversent directement la couche plexiforme interne et une fois arrivées au niveau des spongioblastes, elles se décomposent en une arborisation terminale à branches courtes et variqueuses qui se mettent en contact avec le soma et la tige descendante de ces derniers éléments (b). Chez les oiseaux ces branches terminales se connexionnent d’une façon spéciale avec une catégorie particulière de neurones que nous avons appelés spongioblastes d’association (b). Ces singuliers corpuscules qui possèdent comme des neurones ordinaires un légitime cylindre axe, reçoivent l’impulsion nerveuse centrifuge par leur corps et leurs courtes dendrites en le propageant horizontalement, d’abord et au moyen de l’axon, à des groupes de cellules amacrines siégeant à de grandes distances (d) et ensuite par l’intermédiaire de ces dernières aux articulations interneuronales de la couche plexiforme interne, constituées comme on sait par les panaches descendants des cellules bipolaires d’une part et les dendrites des corpuscules ganglionnaires d’autre part (e).
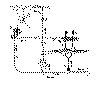
Fig. 6.
Comme spécimen des fibres centrifuges des organes centraux, nous reproduisons dans la figure 7 a, e celles qui se terminent dans le noyau sensitif du thalame. Notez d’abord que ce noyau (noyau latéral de Kœlliker, noyau ventral de Nissl) représente la station où se mettent en rapport les deux neurones sensitifs encéphaliques, savoir, le neurone inférieur formé par les axons du ruban de Reil ou lemnisque interne (fig. 7G), et le neurone supérieur ou thalame-cortical (fig. 7 d) dont le corps siège dans le dit noyau, tandis que le prolongement nerveux, après avoir gagné le corps strié, se termine par des ramifications compliquées, dans la région sensitivo-motrice de l’écorce, s’y mettant en rapport avec les pyramides appartenant à la 3e couche cérébrale (fig. 7 b). Or, mes recherches faites dans le thalame de la souris et du lapin m’ont persuadé qu’un des facteurs, absolument constants, de l’articulation des deux neurones de la voie sensitive supérieure, c’est la ramification terminale des fibres cortico-thalamiques ou descendantes dont l’origine se trouve très probablement dans l’écorce sensitivo-motrice (fig. 7 a, e).

Fig. 7.
Quel est le rôle joué par les fibres centrifuges? Sont-elles des conducteurs destinés à produire dans les articulations des voies sensoriel les un contact très intime qui serait indispensable, ainsi que le pense Duval, pour la bonne propagation de l’impulsion nerveuse? Ou plutôt amèneraient-elles du cerveau quelque modalité d’énergie dont la rapide accumulation dans les stations sensorielles constituerait une condition nécessaire au passage des courants nerveux ascendants? Malheureusement, en l’état actuel de la science il est impossible de faire à ces questions une réponse satisfaisante et categorique.
Les données et les conclusions précédentes se rapportent de préférence à la morphologie de la cellule nerveuse. Dans ce domaine notre notion actuelle basée sur les révélations exactes et précises des méthodes plasmatiques (celles de Golgi et d’Ehrlich) peut être considérée comme définitive. Une fois terminée ou du moins très avancée, l’exploration de la morphologie et des rapports réciproques des neurones, il était tout naturel que les neurologistes portassent leur attention sur le sujet très difilcile et très important de la texture du protoplasma nerveux.
Dans cet intéressant domaine, inauguré très heureusement par Nissl avec la découverte des, grummeaux basophiles du protoplasma, les travaux d’Apathy et de Bethe sur les neurofibrilles ont jeté une vive lumière. Les méthodes imaginées par ces savants étant malheureusement très inconstantes et diffficiles, on a beaucoup travaillé pour en trouver d’autres plus parfaites et abordables. Ces recherches techniques ont abouti à la création des procédés neurofibrillaires de Simarro, de nous-même, de Bielschowsky, Donaggio, Lugaro, etc.
Toutes ces nouvelles ressources techniques ont des avantages dans certains cas particuliers. Notre procédé au nitrate d’argent réduit, sans être supérieur à ceux de Donaggio et de Bielschowsky à l’égard de la différenciation du réticulum protoplasmique adulte, possède quelques avantages, tels que celui de donner de bons résultats chez l’homme en états normal et pathologique, celui de pouvoir s’appliquer couramment chez les animaux âgés de quelques jours, ainsi que dans les organes nerveux en voie de dégénérescence et de régénération, et enfin celui de servir tout particulièrement aux recherches mor phologiques à cause de son efficacité sur des coupes très épaisses et transparentes.
Grâce à ces propriétés d’une méthode qui dans les mains de van Gehuchten, Michotte, G. Sala, Azoulay, Nageotte, Dogiel, Marinesco, Perroncito, Lugaro, Tello etc. a donné de très brillants résultats, nous avons pu ajouter à nos connaissances de l’anatomie et de la physiologie neuronale les acquisitions suivantes:
a. La charpente neurofibrillaire des neurones des vertébrés n’est pas composée, comme l’avait supposé Bethe, par le mélange et l’entrecroisement d’un grand nombre de conducteurs indépendants, mais, au contraire, d’un réseau continu dans lequel apparaissent différenciées certaines travées longues et grosses (les filaments primaires) et d’autres travées courtes, minces et pâles (les filaments secondaires). C’est là une disposition observée aussi ces derniers temps par Donaggio, van Gehuchten, Marinesco, Retzius, Tello, v. Lenhossék, Dogiel et d’autres encore (fig. 8, a, b).
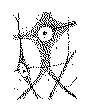
Fig. 8.
b. On rencontre la même disposition réticulaire dans les varicosités et les grossissements des branches nerveuses dans les plaques motrices (nous, Tello), les terminaisons sensitives (Dogiel, Tello), les excroissances des fibres moussues (nous), et les boules terminales des fibres nerveuses en voie de régénération (nous, Perroncito, Marinesco, Nageotte, etc.) Voyez dans la figure 9 c l’aspect du réticulum neurofibrillaire dans les branches terminales d’une plaque motrice, et remarquez que les ramuscules les plus délicats contiennent souvent une anse filamenteuse ou un petit bouton formé par deux ou trois mailles pleines de neuroplasma.
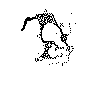
Fig. 9.
L’importance de ces observations, très faciles à contrôler, n’échappera à personne, car si, comme nous le pensons, dans certains cas les neurofibrilles ne se terminent réellement dans le bout libre des branches nerveuses, l’opinion de Bethe et d’Apathy qui considèrent ces filaments comme les seuls organes conducteurs du protoplasma nerveux, ne peut être admise.
c. Le réticulum neurofibrillaire n’est pas un système de conducteurs fixes, stables, mais une charpente capable de subir de notables métamorphoses, soit à l’état physiologique (influence de la chaleur et du froid chez les reptiles et chez les jeunes mammifères, etc.), soit sous l’influence de conditions pathologiques. Parmi les changements dûs à une cause pathologique, mentionnons ceux trouvés par nous et Garcia dans les animaux rabiques (hypertrophie des neurofibrilles, coalescence des faisceaux neurofibrillaires, réabsorption de travées secondaires, etc) confirmés par Marinesco et Franca; ceux très curieux observés chez les animaux refroidis (nous, Tello, Marinesco, Donaggio) et, enfin, ceux rencontrés dans le réticulum des boutons terminaux des fibres nerveuses en voie de régénérescence, et dans le bout périphérique des axons ayant perdu la continuité avec le neurone d’origine.
Nous présentons dans les figures 10, 11 et 12 quelques exemples de ces singulières métamorphoses de la charpente protoplasmique. Notez, dans la figure 10 A les profondes modifications qui se sont opérées dans les neurones moteurs du lézard à la suite de l. Dans les cellules motrices à grande taille (A), les neurofibrilles se sont d’abord approchées en constituant des faisceaux très serrés qui deviennent progressivement, en vertu d’un phénomène de fusion, des cordons absolument homogènes, tandis que dans les petites cellules nerveuses la transformation s’est réduite à l’accumulation dans certains endroits neurofibrillaires de la matière argenthophile (fig. 10 a). Des phénomènes tout à fait semblables sont présentés dans la figure 11 qui reproduit les altérations du réticulum chez le chien rabique (moelle épinière), et dans la figure 12 B, C, dans laquelle nous montrons les très intéressants changements survenus dans les neurones funiculaires du lapin âgé de 15 jours à la suite de l’action, durant 6 heures, d’une température de 10°.

Fig. 10.
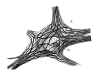
Fig. 11.

Fig. 12.
Enfin la figure 13 démontre les transformations des neurofibrilles dans le bout central d’un nerf écrasé, deux jours après l’opération. On y voit, dans B, les neurofibrilles en voie de régénération pénétrer dans l’intérieur du segment nécrotique de l’axon où elles se terminent par des anses libres; et en C, une portion mortifiée du cylindre axe, envahi par des neurofibrilles ramifiées et en voie d’accroissement. En E, ces derniers filaments, provenant du bourgeonnement d’un axon irrité par le traumatisme, décrivent des tours hélicoïdaux autour de ce dernier qui offre aussi des effilochements et des vacuoles longitudinales. Tous ces singuliers phénomènes, outre qu’ils attestent certaine autonomie des neurofibrilles, montrent aussi la grande capacité qu’elles possèdent de se transformer, en construisant des figures remarquablement variées et variables.

Fig. 13.
En ce qui concerne l’ordre morphologique, nos récentes recherches avec le procédé de l’argent réduit ont révélé:
d. L’existence dans des neurones du grand sympathique humain, d’une espèce particulière de dendrites, les courtes dendrites, caractérisées, outre leur minceur, par le fait qu’elles se distribuent exclusivement par l’espace subcapsulaire (figure 14 b). Ce sont les ramifications terminales de ces appendices qui se mettent en contact avec les nids nerveux qui chez l’homme atteignent, comme l’on peut voir dans la figure 15, une complication extraordinaire.

Fig. 14.

Fig. 15.
c. La présence dans ces mêmes ganglions sympathiques de l’homme de glomérules ou plaques de connexion, c’est-à-dire de plexus spéciaux, formés par la concurrence d’un grand nombre de dendrites appartenant à des neurones différents et dans lesquels viennent s’arboriser et se terminer des fibres nerveuses afférentes.
d. La trouvaille, dans les ganglions sensitifs, de cellules dont le protoplasma est disposé partiellement en un système de cordons anastomotiques (cellules fenêtrées) (fig. 16).
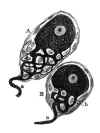
Fig. 16.
e. La présence dans certains neurones sensitifs sympathiques et même cérébelleux d’appendices spéciaux terminés par des boules ou boutons capsulés (fig. 17 a, b). Nous reviendrons ci-après sur l’interprétation de ces bizarres formations qui démontrent la capacité qu’ont les neurones de produire, en vertu de conditions normales et pathologiques, des expansions nouvelles.

Fig. 17.
f. Enfin l’existence dans le bout final en voie d’accroissement des axons embryonnaires, ainsi que dans celui des cylindres axes adultes en cours de régénération, de boutons ou de sphères libres possédant une charpente neurofibrillaire réticulée, etc.
De tout l’ensemble de ces faits découle comme un postulat inévitable la doctrine neuronale de His et de Forel, acceptée par un grand nombre de neurologistes et de physiologistes. Néanmoins —il faut le dire—, quelques unes des inductions physiologiques tirées des observations faites au moyen des méthodes électives pendant ces vingt dernières années ont été combattues et ne peuvent naturellement être considérées comme des dogmes intangibles. La science actuelle, malgré le bien fondé de ses conclusions, n’a pas le droit d’hypothéquer l’avenir. Nos affirmations ne sauraient aller au-delà des révélations des méthodes contemporaines. Peut-être la technique découvrira-t-elle avec le temps quelque procédé de coloration capable de déceler des connexions nouvelles et plus intimes entre les neurones supposés en contact. On ne peut rejeter a priori la possibilité que la forêt inextricable du cerveau dont nous nous imaginons avoir déterminé les dernières branches et feuilles ne possède encore quelque énigmatique système de filaments reliant l’ensemble neuronal comme les lianes rattachent les arbres des forêts tropicales. C’est là une idée qui, pour se présenter à nous avec le prestige de l’unité et de la simplicité, a toujours exercé et exerce encore une puissante suggestion même sur les esprits les plus sereins. Certes, il serait très commode et très économique au point de vue de l’effort analytique que tous les centres nerveux fussent constitués d’un réseau continu intermédiaire entre les nerfs moteurs et les nerfs sensitifs et sensoriels. Malheureusement, la nature semble ignorer notre besoin intellectuel de commodité et d’unité, et se complaît bien souvent dans la complication et la diversité.
D’ailleurs, nous croyons que nous n’avons pas de motif pour être sceptiques. Tout en attendant l’œuvre de l’avenir, restons tranquilles et confiants dans l’avenir de notre œuvre. Rappelons que ces dispositions terminales, que la neurologie moderne a découvertes dans les axons, ont été établies par les révélations concordantes de plusieurs méthodes, et si la science future nous réserve de grandes surprises et des conquêtes admirables il faut penser qu’elle complètera et développera indéfiniment nos connaissances, tout en partant des faits actuels.
L’irrésistible suggestion de la conception réticulaire, dont je vous ai parlé (et laquelle change de formule tous les cinq ou six ans), a condui quelques physiologistes et zooloques à faire des objections à la doctrine de la propagation des courants nerveux par contact ou à distance. Toutes leurs allégations se basent sur les révélations de méthodes incomplètes, infiniment moins démonstratives que celles qui ont servi à bâtir l’imposant édifice de la conception neuronale. Ces arguments appartiennent les uns à l’ordre morphologique, les autres à l’ordre histogénique.
A l’égard des objections morphologiques (desquelles après la découverte de la méthode de Donaggio et de la nôtre on ne parle pas à présent autant qu’avant), je déclarerai seulement que malgré les peines que je me suis données pour apercevoir les prétendues anastomoses intercellulaires dans les préparations faites avec divers procédés de coloration (ceux de Bethe, Simarro, Donaggio, le nôtre, celui de Bielschowsky, etc.), je n’ai jamais réussi à en trouver d’incontestables (c’est-à-dire se montrant aussi claires et nettes que les terminaisons libres), ni dans les plexus nerveux péricellulaires, ni dans les boutons de Held-Auerbach, ni entre les neurofibrilles appartenant à des neurones différents. Et du même avis sont des neurologistes aussi experts et sagaces que His, Kœlliker, Retzius, v. Lenhossék, Duval, van Gehuchten, Lugaro, Schiefferdecker, Déjerine, etc. Si les dites unions intercellulaires ne sont pas le résultat d’une illusion, elles représenteraient des dispositions accidentelles, peut-être monstrueuses dont la valeur serait presque nulle devant la quantité presque infinie des faits de terminaison libre parfaitement observés.
Quant aux arguments histogéniques sur lesquels les adversaires de la doctrine neuronale ont beaucoup insisté ces derniers temps, en les considérant comme les plus graves et les plus décisifs que l’on puisse opposer à la conception neuronale, je répondrai que mes récentes recherches ainsi que celles de Perroncito, Marinesco, Lugaro et Nageotte, faites à l’aide d’un procédé plus expressif que ceux employés par les antineuronistes, prouvent, de la façon la plus péremptoire, le manque de fondement de l’hypothèse du développement discontinu des fibres nerveuses. Purpura, un élève de mon illustre confrère C. Golgi, en analysant avec le procédé du chromate d’argent le processus régénératif des nerfs sectionnés, et tout dernièrement Krassin, de Saint-Pétersbourg, en se servant pour le même objet de la méthode d’Ehrlich, sont arrivés aux mêmes conclusions.
Vous permettrez que j’insiste un peu sur ce point de la régénération et de l’histogénèse normale des nerfs, car c’est une question d’actualité à laquelle nous avons consacré deux années de recherches. D’ailleurs, les conclusions découlant de nos observations —outre leur portée critique— révèlent de très curieux phénomènes de la physiologie du réticulum.
Preuves de la doctrine neurogénétique de Kupffer et His
Vous n’ignorez pas que ces derniers temps on a ressuscité l’ancienne et presque obliée conjecture de Beard et Dohrn sur le mécanisme histogénique des cordons nerveux de l’embryon. C’est la théorie caténaire en vertu de laquelle on admet que les axons des nerfs, au lieu d’être le résultat de l’accroissement de l’expansion primordiale du neuroblaste de His, se formeraient à la suite de la fusion d’un grand nombre de corpuscules ectodermiques émigrés vers la périphérie. Ces éléments reliés en chaîne seraient le siège d’une différenciation fibrillaire d’abord discontinue, puis continue, laquelle aboutirait à la construction d’un grand nombre d’axons ultérieurement fusionnés avec les neurones médullaires. Quant au noyau et au reste du protoplasma non transformé, ils deviendraient dans l’adulte les corpuscules de Schwann.
Voilà la conception qui, avec des variations et même des contradictions que nous n’avons pas le temps d’exposer en détail, a été défendue dernièrement par Sedgwick, Bethe, Joris, Capobianco et Fragnito, Besta, Pighione, etc., à la suite des observations sur l’histogénèse des nerfs chez l’embryon, et par Büngner, Ballance, Bethe, Levi, Durante, van Gehuchten, etc, se basant sur des expériences concernant la régénération des nerfs.
Comme bien des erreurs scientifiques professées de bonne foi par des savants de mérite, la conjecture caténaire est le fruit de deux conditions: l’une subjective, l’autre objective. La première, c’est la tendance fâcheuse mais inévitable pour certains esprits impatients de rejeter l’emploi des méthodes très électives mais se prêtant mal à l’improvisation, telles que celles de Golgi et d’Ehrlich; la seconde, c’est l’application exclusive des procédés simples et commodes mais dépourvus d’action élective sur les axons, et par conséquent incapables de présenter nettement les expansions neuronales et leurs ramifications périphériques.
Pour évitér de regrettables mécomptes dans lesquels tant d’observateurs de talent sont tombés, nous avons choisi (ainsi que Medea, Perroncito, Marinesco et Lugaro) la méthode de l’argent reduit qui a la propriété d’imprégner en une nuance transparente aussi bien les fibres médullées, que celles dépourvues de myéline et en voie de formation.
Or les résultats obtenus démontrent jusqu’à la plus grande évidence qu’à ancun moment de l’évolution des axons on ne surprend de chaînes cellulaires ni de cylindres axes discontinus supposés par les antineuronistes; au contraire, et d’accord avec la doctrine de His et de Kœlliker, aussi bien chez les embryons que dans les nerfs en voie de régénération, les nouvelles fibres se produisent à la suite du bourgeonnement des axons et sont en parfaite continuation avec les neurones moteurs ou sensitifs. Les prétendus neuroblastes périphériques disposés en chaîne représentent des formations tardives, toujours étrangères au protoplasma des prolongements cylindreaxiles; ils appartiennent vraisemblablement au mésoderme. C’est peut-être en vertu d’une action attractive exercée par les axons en voie d’émigrer vers la périphérie que ces éléments, d’abord indifférents, s’approcheraient des fibres et deviendraient les corpuscules de Schwann.
Les preuves de cette doctrine sur l’histogénèse des nerfs se trouvent en abondance aussi bien dans l’histoire de la neurogénèse que dans celle des nerfs régénérés après la section. Mentionnons-en ici quelques-unes.
Preuves tirées du mécanisme régénératif des nerfs
1. Lorsqu’on coupe le nerf sciatique d’un mammifère (lapin, chien ou chat âgés de quelques semaines) et que l’on sacrifie l’animal après une survie de trois jours, on constate, dans les préparations faites par notre procédé, qu’un grand nombre de fibres nerveuses du bout central sont le siège d’un phénomène très actif de bourgeonnement. En poursuivant les fibres vers leur origine on remarque que chacune d’entre elles offre deux portions continues et bien différenciées (fig. 18): le segment ancien tout à fait normal et bien reconnaissable à sa gaine de myéline (F), et le segment néoformé plus mince et pâle et dépourvu de myéline (B). Cette dernière portion de l’axon, dont l’aspect rappelle les fibres de Remak, pénètre dans l’épaisseur de la cicatrice ou au milieu de l’exsudat inflammatoire, se divise très souvent à angle aigu, et ses branches (ou le tronc dans les axons non divisés) finissent au moyen d’une grosse excroissance souvent disposée en forme de bouton (fig. 18 C). Cette boule terminale, parfois fort volumineuse et irrégulière, apparaît d’abord nue; mais dans les jours suivants (depuis le 4e au 6e) elle se montre entourée d’une capsule conjonctive parsemée de noyaux. Quelques branches de bifurcation si tuées au voisinage du tissu cicatriciel émettent des filaments nerveux courbés en arc qui marchent dans l’intérieur du bout central en direction rétrograde (fig. 18 e).

Fig. 18.
Nous avions très bien constaté ces phénomènes de bourgeonnement du bout central depuis le 5e jour de l’opération. Mais Perroncito a eu la fortune de démontrer, en s’aidant du procédé de l’argent réduit, que ce processus de régénérescence est très précoce, apparaissant dès le commencement du 2e jour. Dans nos préparations, toutefois, la formation des boutons et la pénétration de ceux-ci dans le tissu conjonctif intercalaire ne semble commencer qu’à la fin du second jour ou au début du 3e. Nous avons aussi constaté la réalité d’un très curieux phénomène d’effilochement et de dispersion des neurofibrilles des axons, phénomène aboutissant à la création précoce d’un faisceau de fibres nerveuses jeunes dont quelques-unes, en accroissant et en ce ramifiant très activement, donnent lieu à un système de filaments spiraux qui entourent des faisceaux neurofibrillaires rectilignes et axiaux sans abandonner l’espace limité par la gaine de Schwann. Ces singuliers appareils fibrillaires, que nous appellerons organes de Perroncito, manquent souvent chez les jeunes animaux (fig.19B, C). Cependant en étudiant le phénomène régénératif chez le chien et le chat adultes on les observe parfois très abondants surtout dans les gros axons ayant subi des contusions. Jamais on ne peut les surprendre dans les fibres de Remak ou dans les tubes médullaires minces. C’est pourquoi nous sommes incliné à considérer les appareils de Perroncito comme une néoformation pathologique, laquelle conduirait parfois à la production de fibres nerveuses destinées a la cicatrice, mais qui amènerait plus fréquemment la construction de pelotons et de dispositions hélicoïdales infécondes (appareils spiraux du bout central déjà vus par les anciens auteurs, tels que Ranvier, Vanlair, Stroebe, etc).
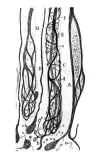
Fig. 19.
Il est aisé de comprendre que le phénomène de Perroncito ainsi que le bourgeonnement précoce des fibres nerveuses du bout central, et surtout la présence des boules d’accroissement dans les extrémités des axons pénétrant dans la cicatrice sont des faits d’une grande valeur en faveur de la doctrine neurogénétique de His, parce que toutes ces dispositions sont antérieures a la phase de multiplication des cellules de Schwann, et par conséquent au processus de différenciation des bandes de Büngner.
2. Quand, en répétant l’expérience de Vulpian, Brown-Séquard, Bethe et van Gehuchten, après la coupure d’un nerf on crée des obstacles entre les deux fragments afin d’en empêcher la réunion immédiate, on constate souvent (pas toujours) trois ou quatre mois après l’opération, une régénération très avancée du segment périphérique. Dans ces circonstances, il en résulte parfois, impossible à déceler à l’examen macroscopique, l’existence d’un cordon continu reliant les deux bouts du nerf interrompu. Voilà le grand argument histologique de l’école caténaire qui deviendrait irréfutable si les antineuronistes eussent soumis la cicatrice intermédiaire à une analyse micrographique approfondie. Malheureusement cette exploration n’a été faite que très incomplètement et à l’aide de méthodes incapables de colorer les fibres nerveuses dépourvues de myéline. Or ce tissu intercalaire supposé vide de fibres néoformées en contient un nombre considérable, comme Marinesco et nous l’avons démontré; seulement les fibres nerveuses ne forment guère un cordon, mais un plexus très lâche, éparpillé par les interstices musculaires et aponeurotiques, et reliant à travers tous les obstacles et souvent après avoir tracé de très grands détours le bout central avec le bout périphérique.
3. Les fibres nerveuses arrivées à destination, c’est-à-dire aux approches du segment périphérique, se divisent à plusieurs reprises en produisant des bouquets nerveux très compliqués déjà signalés par les anciens anatomo-pa thologistes et tout récemment par Purpura, Perroncito, Marinesco et Krassin. Ces branches secondaires abordent le dit segment, en y marchant tantôt par l’intérieur des bandes de Büngner, tantôt par les interstices de ces dernières, en se terminant à des niveaux différents au moyen d’un bouton de croissance. Parfois on surprend quelques fibres se divisant encore dans le sein du bout distal, en s’écartant à angle aigu et s’orientant en direction centrifuge (fig. 20).

Fig. 20.
L’existence de toutes ces ramifications issues des fibres provenant de la cicatrice, et surtout la présence à l’extrême de chaque branche d’une boule d’accroissement constamment orientée vers la périphérie (fig. 20 f) sont des phénomènes absolument inconciliables avec l’hypothèse caténaire. Ajoutons encore que les fibres plus délicates se trouvant dans les bandes de Büngner ne se montrent jamais discontinues mais en continuation évidente avec celles provenant de la cicatrice; elles sont par conséquent indépendantes des cellules de Schwann et des amas adipeux des anciens tubes nerveux.
4. Phénomène des boules (fig. 17). Outre le processus régénératif dont nous venons de parler, il y a aussi des actes de néoformation nerveuse spontanée dans les centres nerveux de l’homme adulte qui sont très propres à l’analyse du mécanisme de la régénération. Ainsi que nous l’avons indiqué plus haut, nos récentes recherches sur la structure des ganglions nerveux chez les mammifères de grande taille ont décelé la présence constante d’un certain nombre de corpuscules sensitifs et sympathiques, dont le corps ainsi que le prolongement principal émettent des fibres nerveuses qui se terminent à des distances variables, tantôt sous la capsule, tantôt dans l’épaisseur même de la substance blanche du ganglion, au moyen d’un bouton d’accroissement entouré d’une enveloppe nucléaire.
Avant d’entreprendre nos recherches sur la régénération des nerfs, nous professions l’opinion que ces curieux appendices à boules constituaient des dispositions stables appartenant à quelque catégorie spéciale de corpuscules sensitifs; mais maintenant, après que nous avons rencontré des faits semblables dans le grand sympathique des vieux, dans le cervelet et les ganglions des animaux attaqués de la rage et d’autres maladies infectieuses, enfin dans les nerfs en voie de régénération; après que Nageotte par ses belles études sur les ganglions des tabétiques a relevé, lui aussi, par le procédé de l’argent réduit l’existence d’un grand nombre de neurones sensitifs à boules et de fibres nerveuses néofor mées, nous ne doutons pas que les appendices terminés par des sphères capsulées représentent tout simplement le résultat d’un très intéressant processus de production nerveuse. Ce serait donc un phénomène physiologique dans certaines limites, qui s’exagèrerait à la suite d’influences toxiques ou d’autres conditions.
Nous ne discuterons pas ici les interprétations que la dite découverte suggère; nous ne dirons rien par exemple sur l’ingénieuse hypothèse de la régénération collatérale imaginée par Nageotte; nous ne nous arrêterons pas non plus à déterminer pour le moment si ces cellules à boules lorsqu’elles produisent des branches nerveuses obéissent à un processus irritatif sans finalité et sans congruence, ou si plutôt elles cherchent à rétablir des voies conductrices dégénérés ou grièvement compromises à la suite de la fatigue fonctionelle ou d’actions toxiques. Nous nous bornerons seulement à faire constater ici que les corpuscules à boules constituent une démonstration éclatante de l’autonomie régénératrice des neurones adultes et de leur capacité de produire par simple bourgeonnement et sans le concours des cellules de Schwann des fibres nouvelles.
Preuves tirées de la neurogénèse embryomnaire
Les suivantes données qui viennent confirmer pleinement la conception neurogénétique de Kupfer, His et Kœlliker sont aussi basées sur les révélations du nitrate d’argent réduit, qui imprègne assez constamment (formule à fixation alcoolique) les fibres nerveuses chez l’embryon du poulet depuis la 60e heure de l’incubation. Voici quelques faits très significatifs.
1. Les neuroblastes de His de la moelle embryonnaire du poulet (fig. 21) affectent d’abord, au commencement du 3e jour, la forme classique en poire eventuellement fusiforme avec une grosse expansion s’accroissant vers la périphérie médullaire et un petit soma formé presque exclusivement par le noyau. Les neurofibrilles, les seuls facteurs constitutifs de la cellule attirant le dépôt argentique, s’initient dans le cône d’origine du prolongement primordial et s’étendent sous la forme de faisceau jusqu’au cône d’accroissement dans l’intérieur duquel elles se terminent en pointe de brosse ou de pinceau. Ces observations confirment donc le cône d’accroissement depuis longtemps démontré à l’aide du chromate d’argent par nous, v. Lenhossék et Retzius; mais la comparaison des coupes colorées par les deux procédés d’imprégnation prouve que cette massue terminale contient, outre le dit pinceau neurofibrillaire, une substance plasmatique incolore (fig. 22) disposée en lames et en épines divergentes (2). Il est inutile de faire remarquer que l’individualité neuronale apparaît certaine dans ces préparations dans lesquelles on ne découvre pas la moindre trace de ces chaînes cellulaires ou de ces bizarres colonnes neuroblastiques anastomosées dont parlent Sedgwick Bethe, Fragnito, Joris, Besta, etc.
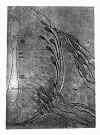
Fig. 21.

Fig. 22.
2. C’est aussi un fait très facile à constater qu’à leurs phases plus embryonnaires (depuis le 2e jour de 1’incubation) les nerfs moteurs et sensitifs ne sont pas composés, ainsi que le soutiennent Beard, Dohrn, Balfour, Fragnito, Levi, etc., par des séries de neuroblastes, mais par des prolongements continus et très minces issus des neurones, lesquels marchent tantôt indépendants, tantôt réunis en faisceaux à travers les vides intercellulaires du mésoderme. Quant aux cellules marginales ou de revêtement (lemmoblastes de Lenhossék), elles apparaîtront plus tardivement (le 4 j ur) siégeant autour des faisceaux nerveux et sans avoir avec ceux-ci que des rapports de contiguité.
3. En examinant dans l’épaisseur des tissus mésodermiques les dernières branches des racines motrices et sensitives, on remarque souvent que chaque fibre se termine librement au moyen d’un bouton d’accroissement tout à fait identique à ceux observés dans les tubes nerveux en voie de dégénération (fig. 23 a). Il est aussi très aisé de surprendre des divisions dichotomiques des jeunes fibres et même des ramifications compliquées (b). Remarquons que dans ces branches périphériques très minces et par conséquent très récentes, on ne trouve jamais de traces de chaînes cellulaires ni même de corpuscules marginaux ou terminaux.
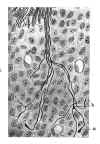
Fig. 23.
4. Les cordons nerveux moteurs examinés dans le voisinage des centres (facial, hypoglose, etc.) manquent complètement de noyaux intérieurs même chez les embryons très avancés (embryon de lapin de 2,5 centimètres). Nous n’insisterons plus sur cette particularité constatée par Kœlliker, His, Gurtwisch, v. Lenhossék, Harrison, etc., et jamais suffisamment expliquée par les adeptes de la théorie caténaire.
5. Ainsi que l’on sait bien, toutes les voies centrales se forment sans le concours des cellules de Schwann, et en vertu d’un processus d’accroissement continu de l’axon des neurones d’association. A ce fait, mis en lumière non seulement par le procédé du nitrate d’argent réduit, mais aussi par les méthodes ordinaires de coloration, nous ajouterons que dans certaines voies, comme celles du cervelet, on trouve souvent, même chez les foetus très développés (chien, chat, lapin) un grand nombre de fibres nerveuses jeunes en voie d’accroissement et pourvues de boutons terminaux.
En résumé: de l’ensemble des observations que nous venons d’exposer et de bien d’autres dont nous n’avons pas le temps de vous entretenir, ressort évidemment comme un postulat inévitable la doctrine neurogénétique de HIS, du savant regretté qui dans les dernières années d’une vie si bien remplie a subi l’injustice de voir qu’une phalange de jeunes observateurs ont qualifié d’erreurs ses plus belles et géniales découvertes.
Je termine en adressant mes salutations très chaleureuses et cordiales à cette docte et sympathique assemblée que je dois aussi remercier infiniment de l’attention et de la bienveillance témoignées durant une aussi longue et fastidieuse conférence.
![]()